

La réponse humorale spécifique commence lorsqu'un lymphocyte B naïf reconnaît son antigène spécifique dans les tissus et les organes lymphoïdes secondaires (voir «Reconnaissance des pathogènes»).
Chaque lymphocyte B a un récepteur d'antigène (BCR: récepteur de cellules B), une immunoglobuline de surface (IgM ou IgD), qui se lie à des domaines spécifiques de l'antigène appelé déterminants antigéniques ou épitopes.

La spécificité du site de liaison antigénique dépend des gènes qui codent les domaines variables (VH et VL); chez les mammifères, la recombinaison et / ou la conversion génique peuvent générer un grand nombre de gènes et, par conséquent, de nombreux BCRs différents, permettant à l'animal de reconnaître presque tous les antigènes externes.
Après reconnaissance, chaque lymphocyte B spécifique est activé, il réalise une prolifération clonale et se différencie en une cellule plasmatique (ou Cellule Sécrétrice d'Anticorps, ASC) capable de synthétiser et sécréter des immunoglobulines (anticorps), spécifiques pour son épitope.
Les cellules T aident à optimiser la réponse humorale spécifique.
Initialement, les cellules B sécrètent des IgM sous forme monomère et avec une spécificité antigénique identique.
En ce qui concerne le type de sécrétion d'anticorps, les antigènes sont classés comme dépendants ou indépendants des cellules T, selon qu'ils ont besoin, ou non, de l'aide des cellules T pour amorcer la réponse des anticorps.
Pour certains antigènes, les cellules dites indépendantes des cellules T (par exemple les lipopolysaccharides bactériens, la flagelline, les polysaccharides pneumococciques, etc.) ne peuvent induire que des anticorps IgM avec une affinité modeste et sans cellules B mémoire.
Presque tous les antigènes externes sont dépendants des cellules T et nécessitent, pour l'activation complète des lymphocytes B, un signal supplémentaire (cytokine) provenant de lymphocytes T-helpers spécifiques qui reconnaissent le même antigène (coopération des cellules T et B).
Dans ce dernier cas, la cellule B agit comme une cellule présentatrice d'antigène, présentant l'antigène traité aux cellules T Helpres qui, lorsqu'elles sont reconnues, permettent le changement isotypique (changement de classe d'anticorps) de l'IgM vers une classe différente d'immunoglobulines ,IgA, IgE ou IgG, en fonction de la Th-cytokine sécrétée, et impliquée dans la stimulation de la cellule B (respectivement IL-4 ou IFNγ). Un autre signal crucial pour la commutation d'anticorps est l'engagement du récepteur CD40 sur les lymphocytes B par CD154 (CD40L).
Sous cette "aide", les cellules B proliférantes subissent une hypermutation somatique de l'immunoglobuline de surface, de sorte qu'elles seront dotées d'un isotype spécifique. Ces lymphocytes B sont ensuite sélectionnés sur la base d'une affinité optimale de l'Ig avec l'antigène lors de leur entrée en contact étroit avec les cellules dendritiques folliculaires qui présentent l'antigène. Seuls les lymphocytes B ayant une affinité d'anticorps élevée pour l'antigène, et capables de la traiter et de le présenter, seront positivement sélectionnés. Par conséquent, seules ces derniers lymphocytes B pourront survivre, proliférer et se différencier en cellules plasmatiques, en synthétisant et sécrétant des anticorps d'une seule classe isotypique avec une spécificité unique et une affinité élevée, en améliorant la capacité à adhérer à l'antigène et, donc, à neutraliser et détruire les agents pathogènes.
Une fois terminé réponse immunitaire primaire, certaines de ces cellules B et les cellules plasmatiques activées et sélectionnées dans ce premier contact, survivront en tant que cellules de mémoire de longue durée de vie, prêtes à être activées dans des expositions ultérieures au même antigène (réponse immunitaire secondaire) en produisant la même classe d'anticorps avec un titre et une affinité supérieurs.
En conclusion, dans la réponse humorale primaire, une seule classe d'IgM est sécrétée initialement sous forme monomère et avec une spécificité antigénique identique. Puis, après quelques jours, en raison de la commutation isotypique, différentes classes d'Ig sont sécrétées, par exemple des IgG. La réponse humorale secondaire induite par une exposition ultérieure au même antigène sera caractérisée principalement par la production d'isotype d'IgG mais avec un titre et une affinité plus élevés (Figure 1).
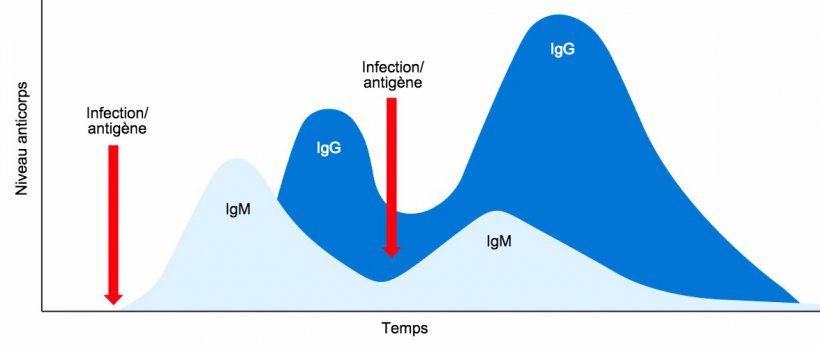
Figure 1: Changement isotypique et réponse immunitaire primaire et secondaire à des expositions ultérieures au même antigène.
Les anticorps peuvent agir contre les pathogènes avec différents mécanismes défensifs:
- Inhibition de l'infectiosité des pathogènes en se liant directement (neutralisation, blocage par adhésion, exclusion immunitaire).
- Activation de la voie alternative (opsonisation) ou classique du complément avec lyse du pathogène.
- En recouvrant l'agent pathogène (ou les cellules infectées), les anticorps permettent que les cellules accessoires, qui reconnaissent des portions du fragment cristallisable (Fc), ingèrent et tuent l'agent pathogène par phagocytose ou opsonisation ou tuent les cellules infectées par ADCC (« Cytotoxicité Cellulaire Dépendante des Anticorps ").
- Régulation de la réponse immunitaire par interaction avec les récepteurs Fc (FcR).
Les FcRs sont exprimées dans de nombreuses cellules effectrices de l'immunité innée, tels que les mastocytes, les neutrophiles, les monocytes, les macrophages et les cellules dendritiques. Plus précisément, les FcyRs (récepteurs Fc pour IgG) sont liée à des immuns complexes, déclenchent l'activation des cellules innées et leur réponse effectrice (phagocytose, stimulation du métabolisme oxydatif, cytotoxicité, libération des peptides vasoactifs et chimiotactiques). Les FcRs des cellules dendritiques favorisent la capture de l'antigène, son traitement et sa présentation aux cellules T cytotoxiques et helpers.
En revanche, la FcR inhibitrices des IgG exprimée dans les cellules B, est impliquée dans la régulation de l'activation et du développement des cellules B et dans la survie des cellules plasmatiques.


